Akronim:
MAXPERSIST
Številka pogodbe:
MN-0023-481
Oddelek:
Oddelek za biologijo
Tip projekta:
Razvojni projekti
Vrsta projekta:
NOO - Načrt za okrevanje in odpornost
Vloga na projektu:
Vodilni
Trajanje:
01.05.2024 - 30.04.2026
Vrednost projekta BF:
178.320,00 €
Vrednost projekta skupaj:
153.628,91 €
Vodja projekta na BF:
Kajin Maja
Projekt poteka v okviru ukrepa C3.K8.IC: »krepitev mednarodne mobilnosti slovenskih raziskovalcev in raziskovalnih organizacij ter za spodbujanje mednarodne vpetosti slovenskih prijaviteljev« v okviru Načrta za okrevanje in odpornost – SKLOP B.
Področje: Populacijska ekologija
Rastlinske in živalske vrste po vsem svetu se soočajo z vedno bolj nepredvidljivimi okoljskimi nihanji. Vedno večja okoljska nihanja pa vplivajo tudi na življanjske cikle organizmov in časovno opredeljene dogodke v življenjskem ciklu, kot je na primer starost ob prvem razmnoževanju. Življenjski cikli se lahko znatno razlikujejo znotraj in med rastlinskimi in živalskimi vrstami. Kombinacije različnih lastnosti življenjskih ciklov (npr. dolgoživa semelparnost, kot pri lososih), so ključne determinante odzivov vrst na okolje. Razlike v življenjskih ciklih med vrstami, populacijami in posamezniki so tako rezultat številnih načinov, na katere je mogoče optimizirati fitnes (angl. fitness). Kdaj v življenju organizma se nek ključni dogodek zgodi (npr. prvo razmnoževanje) je do velike mere pogojeno z genetsko osnovo. Vendar pa okoljski dejavniki močno vplivajo na to, ali se bo nek evolucijsko optimizirani dogodek dejansko zgodil ali ne (torej, ali okolje omogoča, da se prvo razmnoževanje zgodi in kdaj).
Naš planet je zdaj priča večjemu številu sušnih obdobij, požarom, poplavam in drugim ekstremnim klimatskim dogodkom. Globalni podnebni modeli napovedujejo spremembe dveh ključnih okoljskih vidikov: amplitude in nihanja ekstremnih klimatskih dogodkov. Medtem ko naj bi se povprečne vrednosti okoljskih spremenljivk, kot so temperatura in količina padavin na primer, spreminjale, se bo nepredvidljivost v časovnem razporedu vročinskih valov in drugih ključnih dejavnikov ekoloških procesov povečala. Okoljsko stohastičnost opredeljujemo kot pogoji, kjer se spremeni(jo) amplituda in/ali frekvenca okoljskih dejavnikov. V projektu MaxPersist bomo razvili potrebni teoretični napredek za boljše razumevanje, kako se lahko spremenijo lastnosti (in timing) življenjskih ciklov naravnih populacij po vsem svetu, in uporabili to znanje za napovedovanje, kako se lahko populacije stotin živalskih in rastlinskih vrst po vsem svetu obdržijo ob vse večji nepredvidljivosti (t.j. stohastičnosti) okolja.
Splošni cilj projekta:
Projekt preučuje, kako lahko klimatske spremembe – predvsem v luči vedno večje nepredvidljivosti (t.j. povečane stohastičnosti) – potisnejo rastlinske in živalske vrste iz njihovega življenjskega optimuma. To bomo proučevali z demografskimi podatki, t.j. podatki o verjetnosti preživetja in razmnoževanja za vsak starostni (ali velikostni) razred oz. stadij (npr. preživetje ličink, mladih, odraslih osebkov, ipd.). Z velikim številom že obstoječih demografskih podatkov iz javno dostopnih podatkovnih baz (COMADRE [Salguero-Gómez et al. 2016] in COMPADRE [Salguero-Gómez et al. 2015]) ter dodatnih specifično izbranih demografskih podatkih, bomo prikazali uporabnost novo vzpostavljenih teoretičnih dognanj, z namenom uporabe le-teh za napovedovanje usod prostoživečih populacij v stohastičnih okoliščinah.
Delovne hipoteze:
Centralna ideja projekta MaxPersist je bila prvotno osnovana in financirana s strani EU v obliki Marie Curie Actions - Individual Fellowship (MSCA #101032484) v sodelovanju z Univerzo v Oxfordu in SalGo Team (https://www.salgo.ox.ac.uk/welcome). Aktualni projekt NextGenerationEU #NOO MN-0023-481 predstavlja nadaljevanje prvotno zastavljenih idej in omogoča nadaljni razvoj ter aplikacijo novo razvite teorije na velikem številu visoko kvalitetnih demografskih podatkov. Osrednje hipoteze projekta MaxPersist so bile delno že naslovljene, v nadaljevanju projekta pa bodo testirane na še bolj učinkovit način z uporabo novo razvitih teoretičnih dognanj.
H1. Večja okoljska stohastičnost bo vodila do novih oblik življenjskih ciklov. Predvideno povečanje okoljske stohastičnosti bo lahko pripeljalo do pojavljanja alternativnih življenjskih ciklov. To pričakovanje temelji na dokazih, da okoljska stohastičnost pogosto povzroči fenološke spremembe (tj. časovni razpored dogodkov v življenjskem ciklu). Bolj kot se okolje spreminja, več bo spremenjenih časovnih razporedov dogodkov v življenjskem ciklu, kar bo vodilo do nastanka novih alternativ življenjskih ciklov. Poleg tega pričakujemo, da bodo organizmi, katerih življenjski cikel je prilagojen stalnim okoljem, razvili nove strategije za spopadanje s povečano spremenljivostjo okolja – ali pa bodo izumrli.
H2. Večja okoljska stohastičnost bo spremenila kako so demografski parametri (t.j. preživetje ali razmnoževanje pri določeni starosti) povezani med seboj. Teorija napoveduje spremembo v tem, kako so demografski parametri korelirani kot odziv na povečano stohastičnost okolja, ne napoveduje pa smeri te spremembe v korelaciji. Osredotočili se bomo na posledice usmeritve te spremembe korelacij med demografskimi parametri. Predvidevamo, da bo povečana nepredvidljivost okolja povzročila nove selekcijske pritiske tako, da bo vplivala na korelacije med demografskimi parametri. Kot posledica tega, bodo nekatere dolgo obstoječe povezave med demografskimi parametri, kot npr. znana negativna povezava med preživetjem in razmnoževanjem, spremenjene.
H3. Večja okoljska stohastičnost bo oslabila vlogo drugih ekoloških in evolucijskih sil na sposobnost preživetja. Tesno povezane vrste ponavadi kažejo bolj podobne življenjske zgodovine zaradi skupne evolucijske zgodovine. Vendar pričakujemo, da lahko višja amplituda in pogostost okoljskih ekstremov zasenčita učinke filogenetskih in ekoloških omejitev na življenjske zgodovine.
Preliminarni rezultat:
Razvoj nove metode, t.i. S-elastičnost (S-elasticity). S-elastičnost predstavlja biološko pomembno kvantifikacijo tako imenovanega učinka drugega reda na povprečni fitnes osebkov v populaciji. Učinek funkcije drugega reda opisuje kritične točke v dinamičnem obnašanju funkcije, kar pomeni, da identificira točke, kjer funkcija postane nelinearna. V našem primeru je funkcija fitnes. Zato je pomembno vedeti, ali je povprečni fitnes osebkov v neki populaciji za nek demografski parameter konveksne ali konkavne oblike. Konveksnost ali konkavnost fitnesa za določen demografski parameter nam nudi pomembne informacije o tem, ali je potrebna velika sprememba demografskega poarametra, da se fitnes poveča, ali morda samo minimalna. Kljub obstoječi metodi (Caswell 1996) v populacijski ekologiji manjka meritev učinka drugega reda sposobnosti, ki pa je bila računsko zahtevna in je ni bilo enostavno razložiti v biološkem smislu. Na novo opredeljena S-elastičnost je zagotovila potrebno manjkajočo metodo za izpolnitev projektnega širšega cilja.
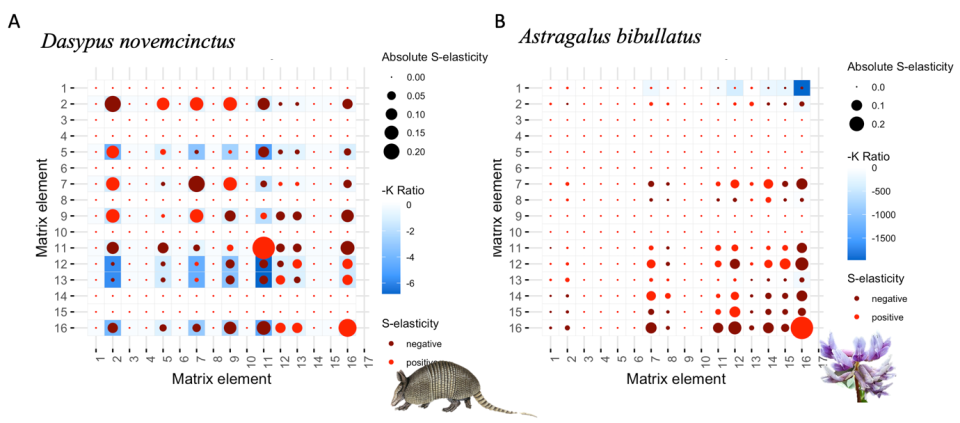
Leganda Slika 1: Prikaz dinamičnih vrednosti S-elastičnosti (svetlo in temn rdeče pike) za živalski (Dasypus novemcinctus) in rastlinski (Astragallus bibullatus) življanjski cikel. Modri kvadrati nakazujejo moč negetivne korelacije med dvema demografskima parametroma, temnejši kot je kvadrat, tem bolj sta dva demografska parametra negativno soodvisna.
Viri:
Caswell, H. 1996. Second Derivatives of Population Growth Rate: Calculation and Applications. Ecology 77:870–879
Salguero-Gómez, R., O. R. Jones, et al. 2015. The COMPADRE Plant Matrix Database: An open online repository for plant demography. Journal of Ecology 103:202–218
Salguero‐Gómez R, Jones OR, et al. 2016. COMADRE: a global data base of animal demography. Journal of Animal Ecology 85(2):371-84